I due saggi primi classificati sono entrambi dedicati ad alcune delle tematiche più care a Gould: le stranezze della natura e il principio dell’utilità a ogni costo. Un altro tema comune ai due saggi è quello dei “vincoli strutturali”, un termine un po’ tecnico e freddo, che Gould non utilizza molto nei suoi saggi. Lo utilizza invece nelle sue pubblicazioni più tecniche, dove dà ad esso anche un significato piuttosto innovativo: un’accezione positiva e non soltanto negativa, laddove i vincoli possono facilitare l’evoluzione, canalizzarla, essere “punti di appoggio” che se da una parte limitano in maniera piuttosto irreversibile i percorsi evolutivi, dall’altra aprono nuove possibilità e campi di esplorazione. Una delle modalità di esplorazione a partire da vincoli strutturali è l’exaptation (Gould & Vrba 1982), che Gould illustra e spiega in maniera eccezionalmente chiara nel saggio sul kiwi.

Figura 1 – La campana della libertà. Sono visibili le due iscrizioni sulla cui diversa origine si sofferma Gould. In basso “Pass and Stow”. In alto “…Proclaim Liberty… By Order Of The Assembly…”.
La campana della libertà
La campana della libertà non mi sembra tra le più riuscite metafore architetturali di Gould, ma, visto che compare nel “best of”, è quella che mi tocca usare! La Liberty Bell si trova a Filadelfia e ha un grande significato storico per gli americani: è forse il simbolo più comunemente associato alla Rivoluzione americana, dato che l’8 luglio 1776 il suo suono radunò i cittadini di Filadelfia per la lettura della Dichiarazione d'indipendenza. Precedentemente, aveva anche suonato per annunciare l’apertura del primo congresso continentale nel 1774, nonché dopo la battaglia di Lexington e Concord nel 1775.
L’attenzione di Gould si focalizza sulla diversa origine delle due iscrizioni (Fig. 1). Gould racconta di aver cercato in tutti i modi di capire che cosa volesse significare “Pass and stow” su questo monumento storico. Dopotutto, la frase impressa nella parte superiore recita: “Proclamerete l’affrancamento nel paese per tutti i suoi abitanti”. Siamo davanti una versione un po’ meno elegante del celebre esempio dei “pennacchi di San Marco” (Gould & Lewontin 1979): in un artefatto, come in una architettura o in un essere vivente, vi sono parti che hanno un’origine funzionale, e altre in cui è molto più preminente la traccia della storia. Per quanto ci sforziamo di trovare un significato, una funzione, “Pass and stow” è soltanto il nome della fonderia che ha prodotto la campana. In altri casi, sono elementi che svolgono una funzione a portare ancora forti i vincoli della storia: è il caso dei pennacchi di San Marco, utilizzati per ospitare i meravigliosi mosaici dei quattro evangelisti ma originatisi per tutt’altre ragioni, strutturali, come sottoprodotti.
L’uovo del kiwi
Il kiwi (pronuncia: kivi) di cui Gould parla nel saggio “L’uovo del kiwi e la campana della libertà” è un uccello con le ali atrofizzate, grande quanto una gallina, diffuso soltanto in Nuova Zelanda. Come da copione gouldiano, la descrizione vivida e accurata del kiwi è seguita da una domanda, una di “quelle” famose domande. La maggiore stranezza del kiwi sta nell’uovo, in particolare nella sua enormità rispetto al corpo: esso può raggiungere il 25% del peso corporeo della femmina, e al massimo stadio di sviluppo si estende dalla parte più alta del petto fino alla cloaca! Per fare un paragone improprio, pensiamo a una donna di 40 kg con un feto pesante 10 kg… La femmina kiwi – che depone due o tre uova per ogni covata, a distanza di circa 33 giorni – deve ancheggiare, con le gambe divaricate, per vari giorni prima della deposizione, esponendosi ai pericoli e nutrendosi con difficoltà.
Ciò che è più interessante per Gould sono le strategie che utilizziamo quando cerchiamo di spiegare la dimensione, evidentemente sproporzionata, di questo uovo. A me è capitato diverse volte, proprio su ispirazione di questo saggio, di sfidare gruppi di persone a trovare una spiegazione (cf. Serrelli 2008). Proprio come previsto da Gould, la strategia più diffusa punta sui vantaggi che un uovo così grande può portare, non tanto alla madre, quanto al nascituro: la quantità di nutrimento, la possibilità di svilupparsi maggiormente prima della schiusa. Ma alcune persone sono più creative, e immaginano uno scenario in cui la dimensione dell’uovo impedisca ad esso di cadere nelle crepe di un ipotetico terreno roccioso in cui i kiwi vivrebbero.
Una spiegazione completamente
differente, e molto più affascinante per Gould, è quella trovata da William
Alexander Calder III (1934-2002): una spiegazione basata sulle regole
strutturali dello sviluppo, non sul valore adattativo di un carattere.
Innanzitutto, bisogna collegare i kiwi con la loro origine evolutiva
(filogenetica) e con i “parenti” da cui discesero: i kiwi sarebbero “nani
filogenetici”, si sarebbero cioè evoluti con un rimpicciolimento da un gruppo
uccelli molto più grossi, assimilabili ai moa. In tale diminuzione di dimensione sarebbe stata conservata, immodificata, una
regola strutturale: il rapporto che regola la proporzione tra peso
dell’individuo e peso dell’uovo. Un vincolo strutturale.
Ogni carattere nell’organismo è
correlato a molti altri. In generale, negli esseri viventi si osserva che una
minore dimensione del corpo non si accompagna a un rimpicciolimento altrettanto
veloce di molti caratteri particolari. Le “curve allometriche” (Fig. 2) sono
descrizioni matematiche di questo fenomeno: potremmo dire che esprimono il
mutamento di proporzioni al diminuire del peso corporeo. In ogni specie di uccelli
esiste una curva allometrica che associa a differenti pesi del corpo un
corrispondente peso dell’uovo. Una regola come questa (intraspecifica) ha senso
e diventa evidente se si mettono in ordine di peso i membri della stessa
popolazione, ed essa descrive il peso delle loro uova. A diverse specie
corrispondono diverse curve allometriche. La curva allometrica INTERspecifica
mostra in armonica disposizione tutte le diverse specie, dal colibrì al moa
passando per le galline, ognuna con uno specifico rapporto (medio) tra corpo e
uovo. Per ogni punto della curva interspecifica passa quella intraspecifica della
specie corrispondente. Insomma, normalmente ogni specie di uccelli ha una curva allometrica intraspecifica adeguata al peso medio del corpo dei suoi membri.
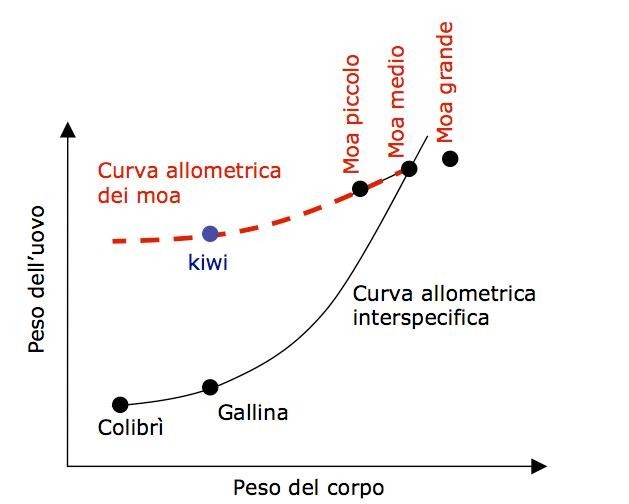
Figura 2 – Una curva allometrica descrive le proporzioni di un tratto al variare della dimensione complessiva dell’organismo. In nero: curva allometrica interspecifica (dal colibrì al moa) che descrive il peso medio dell’uovo di ogni specie. In rosso tratteggiato: curva allometrica intraspecifica dei moa. La curva intraspecifica è meno pendente di quella interspecifica. La sproporzione dell’uovo del kiwi si deve alla conservazione di una regola strutturale attraverso il nanismo.
Ci si aspetterebbe che il kiwi avesse una curva allometrica – e quindi un uovo – simile a quella delle galline domestiche. Invece il suo uovo è sei volte più grande. L’ipotesi di Calder ripresa da Gould è che l’uovo dei kiwi, mentre essi diminuivano di dimensione nell’adattarsi alla vita insulare, sia “sceso” lungo la curva allometrica intraspecifica dei moa (da un adulto moa grande, a un adulto moa piccolo) che è molto meno pendente di quella interspecifica: che in questo processo, sotto pressioni selettive effettivamente molto lievi, non sia avvenuta la modificazione di questa regola strutturale. La grandezza dell’uovo sarebbe quindi nient’altro che un effetto collaterale della diminuzione di dimensione che sottosta ai vincoli strutturali dell’organismo.
E l’utilità? È verosimile che un uovo di grandi dimensioni consenta all’embrione che in esso si sviluppa di disporre di maggiore nutrimento e di nascere più preparato alla vita. Ma, come sottolinea Gould, una spiegazione biologica dovrebbe prima di tutto comprendere i processi strutturali attraverso i quali emerge un carattere, non i motivi (adattativi) per cui esso si conserva! La selezione naturale spiega semmai la conservazione dei caratteri strutturali emersi: la scomodità di un uovo così grande non ha portato all’estinzione del kiwi perché nell’ambiente non vi sono molti predatori – la pressione selettiva è molto debole. La selezione naturale funziona sulla variazione tra gli individui, la quale prende forma attraverso vincoli strutturali che sono difficilmente modificabili: vengono ereditati e sono il punto di partenza per la produzione di ulteriori novità.
Mi fa piacere che i kiwi se la cavino molto bene con le loro grandi uova. Ma possiamo concludere che queste uova smisurate siano state create dalla selezione naturale in vista di questi benefici? Questo assunto – che passa con la più grande facilità dalla funzione corrente di un carattere alla ragione per la sua origine – è, secondo me, l’errore più grave e diffuso presente nella mia professione, giacché questa falsa inferenza è alla base di centinaia di racconti convenzionali sulle vie seguite dall’evoluzione. Vorrei identificare questo errore di ragionamento con una frase che potrebbe essere un motto: L’utilità attuale non può essere messa sullo stesso piano con l’origine storica, ovvero, quando si mostra che qualcosa funziona bene, non si è ancora risolto il problema di come, quando o perché quella tal cosa abbia avuto origine. Io propongo una ragione semplice per etichettare come erronea un’inferenza automatica dall’utilità attuale all’origine storica: una buona funzione può sempre avere un’interpretazione alternativa. Una struttura oggi utile può essere stata costruita dalla selezione naturale per la sua funzione attuale (non nego che spesso l’inferenza regga), ma potrebbe essere stata sviluppata anche per un’altra ragione (o per nessuna ragione funzionale particolare) ed essere stata poi cooptata per il suo uso presente (KIWI91, p. 114).
I casi del secondo e terzo tipo sono “catturati” dal famoso termine, exaptation, introdotto da Gould e Vrba nel 1982 proprio per sopperire a quello che vedevano un “punto cieco” della teoria dell’evoluzione. Nonostante la sua probabile utilità odierna, il grande uovo del kiwi è come “Pass and Stow” sulla Campana: un prodotto dell’inerzia della storia, pronto perché ad esso, se non viene eliminato, si inventi un nuovo uso.
[pagebreak]
Capezzoli maschili e glande clitorideo
…il pregiudizio dell’utilità ha suscitato inutili sofferenze e ansie a milioni di persone (CMGC91, p. 129).
È ciò che si legge nel saggio vincitore, il famoso Tits and clits, tradotto in “Capezzoli maschili e glande clitorideo”. Qui il discorso sui vincoli strutturali è applicato a noi stessi, uomini e donne, e si unisce alla pregnanza sociale della scienza. “Voglio dire, dopo tutto: perché la gente dovrebbe preoccuparsi tanto di idee speculative, anche se sbagliate, che però non procurano danni tangibili alla vita delle persone? […] tutto questo non è rigorosamente entre nous?” (Ivi, p. 134). Domanda retorica da parte di Gould, dato che il suo saggio si occupa proprio delle potenziali ripercussioni delle idee, scientifiche e non, sulle vite delle persone. Il pregiudizio dell’utilità è stato applicato a un carattere, l’orgasmo femminile, determinando ancora una volta un’aspettativa: quella della sua localizzazione anatomica vaginale. Nella teoria dell’evoluzione, il funzionalismo si esprime nei termini di:
…una lotta fra organismi per un successo riproduttivo differenziale. Il piacere sessuale, in sintesi, dovette evolversi come stimolo per la riproduzione. Questa formulazione funziona per il maschio giacché il picco dell’eccitazione sessuale coincide con l’eiaculazione: un concomitante diretto e primario del coito. Per gli uomini il massimo piacere è associato alla massima probabilità di produrre prole. In questa prospettiva, anche il piacere sessuale delle donne dovrebbe essere incentrato sull’atto che causa la fecondazione: ossia sulla copula stessa (Ivi, p. 131).
Ecco che la teoria diviene normativa, e l’orgasmo clitorideo è ridotto a un’anomalia, a una disfunzionalità. Questo è ciò che si ritrova, analizza Gould, nella teoria di Sigmund Freud della transizione dall’orgasmo clitorideo all’orgasmo vaginale, essenza della maturità sessuale femminile:
Questo dogma del passaggio dall’orgasmo clitorideo a quello vaginale divenne luogo comune della cultura popolare […], plasmò le attese di milioni di donne istruite e “illuminate”, le quali si sentirono ripetere da una schiera di psicoanalisti e da centinaia di articoli […] che dovevano compiere questa transizione biologicamente impossibile per raggiungere la maturità sessuale (Ivi, pp. 134-5).
La frigidità e la nevrosi sarebbero state legate alla mancanza o alla difficoltà di questa transizione. Il legame tra orgasmo e copula si ritrova in Freud non per l’influenza diretta di un modello biologico sbagliato (la teoria di Freud è abiologica), bensì per un modo di pensare, quello funzionalistico, più fondamentale. Un modo di pensare così forte da resistere alle evidenze contrarie, ai fatti:
Come le donne sanno fin dall’alba dei tempi, la localizzazione primaria per la stimolazione dell’orgasmo femminile è incentrata sulla clitoride. La rivoluzione scatenata dal rapporto Kinsey del 1953 [seguito da molti altri studi] ha reso oggi disponibile anche a quegli uomini che, per qualsiasi ragione, non ci fossero arrivati da soli […] (Ivi, p. 129).
Si tratta di indagini quantitative sulla sessualità delle donne americane. Il qui citato Alfred Kinsey è un mito per Gould, a lui è dedicato un saggio in Il sorriso del fenicottero.
Nonostante le evidenze contrarie, per molti anni grandi biologi evoluzionisti (Morris 1969, Eibl-Eibesfeldt 1975) hanno dato addirittura per scontato che l’orgasmo femminile sia vaginale, costruendo poi le loro ipotesi di adattamento alla riproduzione e alla monogamia. Inoltre, anche essere avversari dell’associazione tra orgasmo e riproduzione non significa fare a meno del principio di utilità, di adattamento funzionale. Anzi. È il caso di Sarah Hrdy, antropologa e primatologa ora emerita alla UC Davis, citata da Gould come scienziata che sosteneva che…
…la dissociazione tra orgasmo e copula è un adattamento che favorisce il comportamento promiscuo; ciò permetterebbe alla femmina di assicurarsi il sostegno di vari maschi, mettendosi così in condizione di impedire a qualcuno di loro di arrecare danno ai suoi piccoli (Ivi, p. 133, corsivo mio).
Insomma, si dimostra facilmente che spiegazioni funzionali speculative ad hoc si possono trovare per ogni tratto e anche per il suo contrario.
Eppure l’ovvia alternativa non adattiva, quella strutturale, è chiara di fronte a noi come il fatto più elementare della sessualità umana: l’omologia fra il pene e la clitoride (Ivi, p. 137).
La clitoride – spiegava Kinsey e riportava Gould – è altrettanto riccamente dotata di terminazioni nervose del pene, ed è perciò capace di eccitazione. Le pareti della vagina, d’altra parte, sono “prive di organi terminali del tatto e sono del tutto insensibili” e “per la maggior parte delle donne l’insensibilità è estesa a ogni parte della vagina”. Vediamo che questa volta l’omologia esplicativa non è – come nel caso del kiwi – tra parenti, bensì tra i sessi di una specie (Fig. 3). E il medesimo principio che spiega i capezzoli maschili, con i quali concludiamo degnamente la trattazione di questo meritevole primo classificato:
I rapporti Kinsey sono due libri sul comportamento sessuale dell’essere umano: Sexual Behaviour in the Human Male (1948) e Sexual Behaviour in the Human Female (1953), scritti dal biologo Alfred Kinsey e colleghi. I risultati di questi lavori ebbero un’enorme risonanza presso la gente comune e furono immediatamente considerati controversi e sensazionalistici: si occupavano di argomenti che in precedenza erano considerati tabù e sfidavano le conoscenze convenzionali sulla sessualità (ad esempio, la credenza che l’eterosessualità e l’astinenza fossero la norma, sia statisticamente che eticamente).
Supponiamo di prendere l’avvio da un punto di vista diverso, concentrandoci sulle regole di accrescimento e di sviluppo. Le differenze esterne fra maschio e femmina si sviluppano gradualmente da un embrione così generalizzato che in esso non è affatto facile determinare il sesso. La clitoride e il pene sono uno stesso organo, identico nei due sessi agli inizi della vita embrionale, che in seguito si ingrossa nei feti maschili in conseguenza dell’effetto del testosterone. Similmente, le grandi labbra nella donna e i sacchi dello scroto nell’uomo sono la stessa struttura, indistinguibile negli embrioni più giovani, ma che in seguito si ingrandiscono, si ripiegano e si saldano lungo la linea mediana nei feti maschili.
Non ho dubbi sul fatto che la grandezza e la sensibilità delle mammelle femminili debba essere considerata un adattamento nei mammiferi, ma la versione più piccola del maschio non ha bisogno di alcuna spiegazione adattiva. Maschi e femmine non sono entità separate, formate indipendentemente dalla selezione naturale. I due sessi sono variazioni su un singolo piano fondamentale, che si differenziano nella parte posteriore dello sviluppo embrionale. I mammiferi maschi hanno i capezzoli perché le femmine ne hanno bisogno… (CMGC91, p. 128).
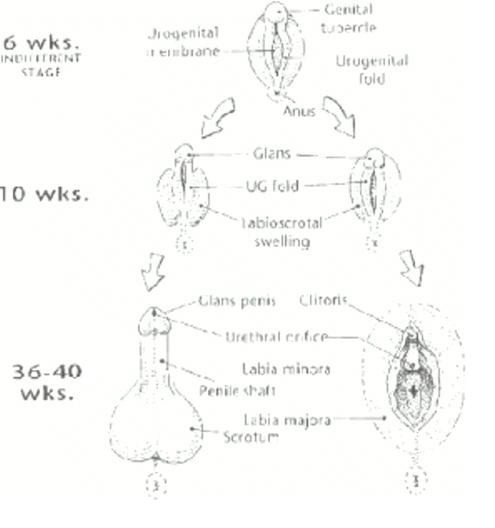
Figura 3 – Omologia tra organi sessuali maschili e femminili,
che libera l’orgasmo femminile dalla necessità di una stretta associazione con
la fitness riproduttiva, e quindi da determinate aspettative sulla sua collocazione
strutturale (che possono, a loro volta, ripercuotersi in termini di criteri di
“normalità”).
Fonte: ML Gustafson, PK Donahoe (1994) Male sex determination:
Current Concepts of Male Sexual Differentiation, Annual Review of Medicine
45:505-524.
